Cross-sectional anatomy of the spinal cord
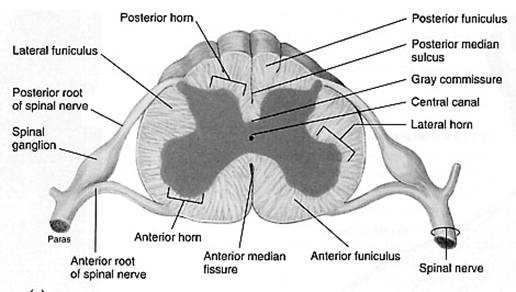
The spinal cord appears to be somewhat flat with two grooves that mark its surface. The two grooves are named as follows: the ventral (anterior) median fissure and the more shallow dorsal (posterior) median sulcus. These two grooves run the length of the cord and partially divide it into right and left halves. The gray matte of the spinal cord is located at its center, with the white matter surrounding it.
Gray Matter and Spinal roots
In a cross section of the spine, the gray matter looks similar to the letter "H". It is two lateral gray masses connected by a gray crossbar referred to as the gray commissure, all of which surrounds the central canal. The two dorsal projections of gray matter are called the dorsal (posterior) horns and the ventral (anterior) horns. These horns form columns of gray matter that run the length of the spinal cord. An additional pair of gray matter columns, the lateral horns, are present in the thoracic and superior lumbar segments of the cord.
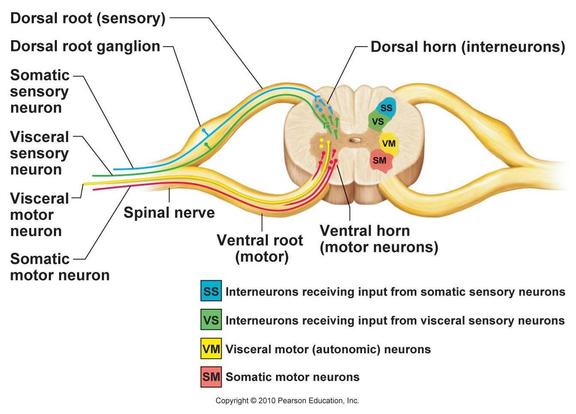
All of the neurons whose cell bodies are located within the spinal cord's gray mater are multipolar. The dorsal horns consist entirely of interneurons and the ventral horns have some interneurons, but are mostly comprised of the cell bodies of somatic motor neurons. These motor neurons send out their axons to skeletal muscles (or effector organs) via the ventral rootlets that fuse to become the ventral roots of the spinal cord.
The amount of ventral gray matter present at a given level of the spinal cord reflects the amount of skeletal muscle innervated at that level. Because of this, the ventral horns are largest in the limb-innervating cervical and lumbar regions of the spinal cord. Also, they are responsible fro the enlargement of the cord seen in those regions.
The lateral horns are autonomic, which is the sympathetic division, motor neurons that serve all of the visceral organs. Their axons leave the cord via the ventral root along with the axons of the somatic motor neurons. Because the ventral roots contain both somatic and autonomic efferents, they serve both motor divisions of the peripheral nervous system.
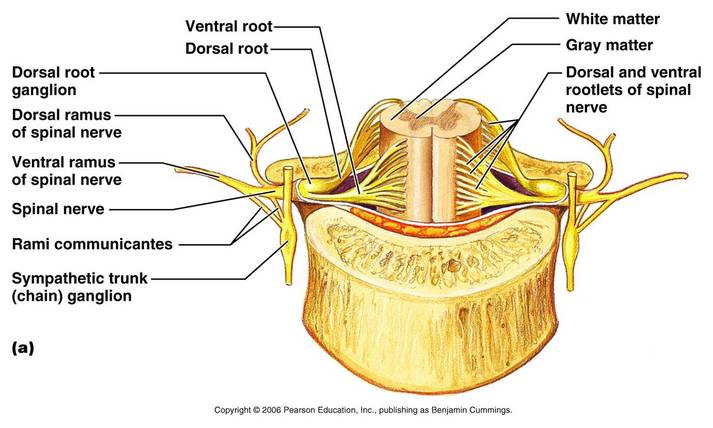
Afferent fibers carrying impulses from the peripheral sensory receptors form the dorsal roots of the spinal cord that fan out as the dorsal rootlets before they enter the spinal cord. The cell bodies of the associated sensory neurons are found in an enlarged region of the dorsal root called the dorsal root ganglion or spinal ganglion. After entering the cord, the axons of these neurons may take a number of routes. Some enter the dorsal white matter of the cord directly and travel to synapse at higher cord or brain levels. Or synapse with interneurons in the dorsal horns of the spinal cord gray matter at their entry-level.
The dorsal and ventral roots are very short and fuse laterally to for m the spinal nerves. The spinal nerves, which are part of the peripheral nervous system, will not be discussed here.
The spinal gray matter can be further divided according to its neurons' relative involvement in the innervation of somatic and visceral regions of the body. There are four zones evident in this gray matter: somatic sensory (SS), visceral (autonomic) sensory (VS), visceral motor (VM), and somatic motor (SM).
All of the neurons whose cell bodies are located within the spinal cord's gray mater are multipolar. The dorsal horns consist entirely of interneurons and the ventral horns have some interneurons, but are mostly comprised of the cell bodies of somatic motor neurons. These motor neurons send out their axons to skeletal muscles (or effector organs) via the ventral rootlets that fuse to become the ventral roots of the spinal cord.
The amount of ventral gray matter present at a given level of the spinal cord reflects the amount of skeletal muscle innervated at that level. Because of this, the ventral horns are largest in the limb-innervating cervical and lumbar regions of the spinal cord. Also, they are responsible fro the enlargement of the cord seen in those regions.
The lateral horns are autonomic, which is the sympathetic division, motor neurons that serve all of the visceral organs. Their axons leave the cord via the ventral root along with the axons of the somatic motor neurons. Because the ventral roots contain both somatic and autonomic efferents, they serve both motor divisions of the peripheral nervous system.
Afferent fibers carrying impulses from the peripheral sensory receptors form the dorsal roots of the spinal cord that fan out as the dorsal rootlets before they enter the spinal cord. The cell bodies of the associated sensory neurons are found in an enlarged region of the dorsal root called the dorsal root ganglion or spinal ganglion. After entering the cord, the axons of these neurons may take a number of routes. Some enter the dorsal white matter of the cord directly and travel to synapse at higher cord or brain levels. Or synapse with interneurons in the dorsal horns of the spinal cord gray matter at their entry-level.
The dorsal and ventral roots are very short and fuse laterally to for m the spinal nerves. The spinal nerves, which are part of the peripheral nervous system, will not be discussed here.
The spinal gray matter can be further divided according to its neurons' relative involvement in the innervation of somatic and visceral regions of the body. There are four zones evident in this gray matter: somatic sensory (SS), visceral (autonomic) sensory (VS), visceral motor (VM), and somatic motor (SM).
White Matter
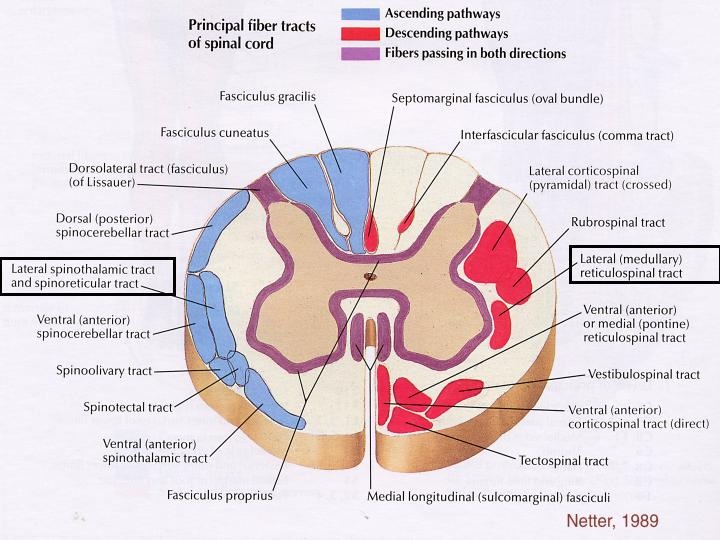
The white matter of the spinal cord is composed of myelinated and unmyelinated nerve fibers that allow communication between different parts of the spinal cord and between the cord and the brain. These fibers run in three directions: ascending (up to higher centers), descending (down into the cord from the brain or within the cord to lower levels), and transversely (across the cord from one side to another). Ascending and descending tracts make up most of the white matter.
The white matter on each side of the cord is divided into three white columns, or funiculi. They are named according to their position as the dorsal (posterior). lateral, and ventral (anterior) funiculi. Each funiculus contains several fiber tracts, where each tract is made up of axons with similar destinations and functions. With a few exceptions the names of the spinal tracts reveal both their origin and destination.
All major spinal tracts are part of multineuron pathways that connect the brain to the body periphery. These great ascending and descending pathways contain not only spinal cord neurons, but also parts of peripheral neurons and neurons in the brain. Before we get specific about individual tracts, here are some generalizations about them and the pathways they contribute to:
1.) Decussation- Most pathways cross from one side of the CNS to the other (decussate) at some point along their journey.
2.) Relay- Most pathways consist of a chain of two or three neurons (a relay) that contribute to successive tracts of the pathway.
3.) Somatotophy- Most pathways exhibit somatotophy, which is a precise spatial relationship between the tract fibers that reflects the orderly mapping of the body.
4.) Symmetry- All pathways and tracts are paired symmetrically, with a member of the pair present on either side of the spinal cord or brain.
The white matter on each side of the cord is divided into three white columns, or funiculi. They are named according to their position as the dorsal (posterior). lateral, and ventral (anterior) funiculi. Each funiculus contains several fiber tracts, where each tract is made up of axons with similar destinations and functions. With a few exceptions the names of the spinal tracts reveal both their origin and destination.
All major spinal tracts are part of multineuron pathways that connect the brain to the body periphery. These great ascending and descending pathways contain not only spinal cord neurons, but also parts of peripheral neurons and neurons in the brain. Before we get specific about individual tracts, here are some generalizations about them and the pathways they contribute to:
1.) Decussation- Most pathways cross from one side of the CNS to the other (decussate) at some point along their journey.
2.) Relay- Most pathways consist of a chain of two or three neurons (a relay) that contribute to successive tracts of the pathway.
3.) Somatotophy- Most pathways exhibit somatotophy, which is a precise spatial relationship between the tract fibers that reflects the orderly mapping of the body.
4.) Symmetry- All pathways and tracts are paired symmetrically, with a member of the pair present on either side of the spinal cord or brain.
Ascending Pathways to the brain
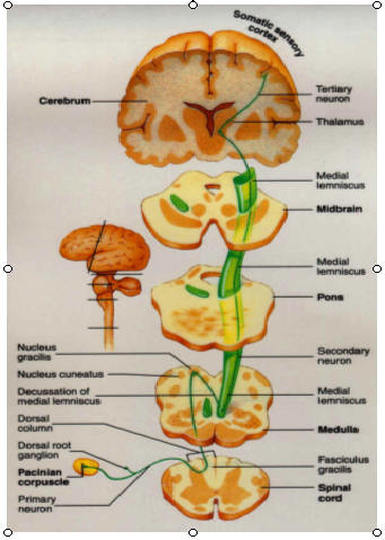
The ascending pathways conduct sensory impulse upward, typically through chains of three successive neurons to various areas of the brain.
1.)First-Order Neuron: cell bodies reside in a ganglion (dorsal root or cranial), conducts impulses from the cutaneous receptors of the skin and from proprioceptors to the spinal cord or brain stem, where they synapse with second order neurons .
2.) Second-Order Neuron: cell bodies reside in the dorsal horn of the spinal cord or in medullary nuclei, and they transmit impulses to the thalamus or cerebellum where they synapse.
3.) Third-Order Neuron: cell bodies in the thalamus. They relay impulses to the somatosensory cortex of the cerebrum.
In general, somatosensory information is conveyed along three main pathways on each side of the spinal cord. Two of these pathways, the dorsal column-medial lemniscal and the spinothalamic, transmit impulses via the thalamus to the sensory cortex for conscious interpretation. Collectively the inputs of these sister tracts provide discriminative touch and conscious proprioception. Both pathways decussate-- the first in the medulla and the second in the spinal cord.
The third pathway, the spinocerebellar pathway, terminates in the cerebellum and does not contribute to sensory perception.
1.)Dorsal column-medial lemniscal pathways: The dorsal column-medial lemniscal pathways mediate precise, straight-through transmission of inputs from a single type (or few related types) of sensory receptor that can be localized on the body surface, such as discriminative touch and vibrations. These pathways are formed by paired tracts of the dorsal white column of the spinal cord (fasciculus cuneatus and fasciculus gracilis) and the medial lemniscus. The medial lemniscus arises in the medulla and terminates in the thalamus. Impulses are forwarded to specific areas of the somatosensory cortex.
2.) Anterolateral pathways: The anterolateral pathways receive input from many different types of sensory receptors and make multiple synapses in the brain stem. These pathways are largely formed by the lateral and ventral spinothalamic tracts. Their fibers cross over in the spinal cord. Most of the fibers in these pathways transmit pain, temperature, and coarse touch impulses, sensations that we are aware of, but are difficult to localize precisely on the body surface.
3.) Spinocerebellar tracts: The last pair of ascending pathways are the ventral and dorsal spinocerebellar tracts. They convey information about muscle or tendon stretch to the cerebellum, which uses this information to coordinate skeletal muscle activity. these pathways do not contribute to conscious sensation. The fibers of the spinocerebellar pathways either do not decussate or else cross over twice.
1.)First-Order Neuron: cell bodies reside in a ganglion (dorsal root or cranial), conducts impulses from the cutaneous receptors of the skin and from proprioceptors to the spinal cord or brain stem, where they synapse with second order neurons .
2.) Second-Order Neuron: cell bodies reside in the dorsal horn of the spinal cord or in medullary nuclei, and they transmit impulses to the thalamus or cerebellum where they synapse.
3.) Third-Order Neuron: cell bodies in the thalamus. They relay impulses to the somatosensory cortex of the cerebrum.
In general, somatosensory information is conveyed along three main pathways on each side of the spinal cord. Two of these pathways, the dorsal column-medial lemniscal and the spinothalamic, transmit impulses via the thalamus to the sensory cortex for conscious interpretation. Collectively the inputs of these sister tracts provide discriminative touch and conscious proprioception. Both pathways decussate-- the first in the medulla and the second in the spinal cord.
The third pathway, the spinocerebellar pathway, terminates in the cerebellum and does not contribute to sensory perception.
1.)Dorsal column-medial lemniscal pathways: The dorsal column-medial lemniscal pathways mediate precise, straight-through transmission of inputs from a single type (or few related types) of sensory receptor that can be localized on the body surface, such as discriminative touch and vibrations. These pathways are formed by paired tracts of the dorsal white column of the spinal cord (fasciculus cuneatus and fasciculus gracilis) and the medial lemniscus. The medial lemniscus arises in the medulla and terminates in the thalamus. Impulses are forwarded to specific areas of the somatosensory cortex.
2.) Anterolateral pathways: The anterolateral pathways receive input from many different types of sensory receptors and make multiple synapses in the brain stem. These pathways are largely formed by the lateral and ventral spinothalamic tracts. Their fibers cross over in the spinal cord. Most of the fibers in these pathways transmit pain, temperature, and coarse touch impulses, sensations that we are aware of, but are difficult to localize precisely on the body surface.
3.) Spinocerebellar tracts: The last pair of ascending pathways are the ventral and dorsal spinocerebellar tracts. They convey information about muscle or tendon stretch to the cerebellum, which uses this information to coordinate skeletal muscle activity. these pathways do not contribute to conscious sensation. The fibers of the spinocerebellar pathways either do not decussate or else cross over twice.
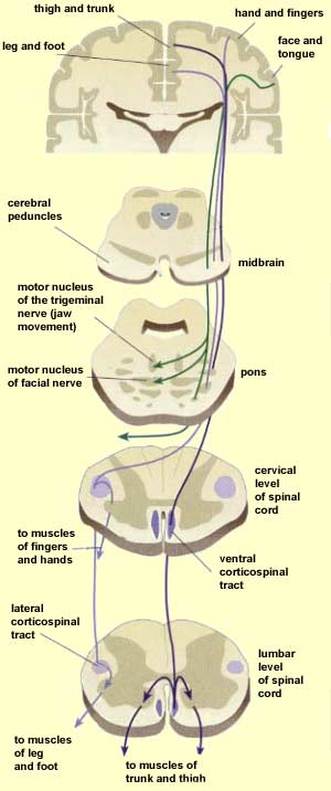
Descending Pathways and Tracts
The descending tracts that deliver efferent impulses from the brain to the spinal cord are divided into two groups: the direct pathways (equivalent to the pyramidal tracts) and the indirect pathways (all others). Motor pathways involve two neurons, referred to as the upper and lower motor neurons. The pyramidal cells of the motor cortex, as well as the neurons in subcortical motor nuclei that give rise to other descending motor pathways, are called upper motor neurons. The ventral horn motor neurons, which actually innervate the skeletal muscles, are called lower motor neurons.
1.) The Direct (Pyramidal) System: The direct pathways originate mainly with the pyramidal neurons located in the precentral gyri. These neurons send impulses through the brain stem via the pyramidal (corticospinal) tracts. The direct pathways are so named because their axons descend without synapsing from the pyramidal neurons to the spinal cord. There they synapse either with interneurons or with ventral horn motor neurons. Stimulations of the ventral horn neurons activates the skeletal muscles with which they are associated, The direct pathway primarily regulates fast and fine movements such as writing.
2.) The Indirect (Extrapyramidal) System: The indirect system includes brain stem motor nuclei and all motor pathways except the pyramidal pathways. These tracts were formerly lumped together as the extrapyramidal system because their nuclei of origin were presumed to be independent of the pyramidal tracts. This term is still widely used clinically. However, pyramidal tract neurons are now known to project to and influence the activity of most "extrapyramidal" nuclei, so modern anatomists prefer to use the term indirect, or multineuronal pathways.
These motor pathways are complex and multisynaptic. They are most involved in regulating the axial muscles that maintain balance and posture, the muscles controlling coarse limb movements, and head, neck, and eye movements that follow objects in the visual field. Many of the activities controlled by subcortical motor nuclei depend heavily on reflex activity.
Overall, the reticulospinal and vestibulospinal tracts maintain balance by varying tone of postural muscles. The rubrospinal tracts control flexor muscles, whereas the tectospinal tracts and the superior colliculi mediate head movements in response to visual stimuli.
1.) The Direct (Pyramidal) System: The direct pathways originate mainly with the pyramidal neurons located in the precentral gyri. These neurons send impulses through the brain stem via the pyramidal (corticospinal) tracts. The direct pathways are so named because their axons descend without synapsing from the pyramidal neurons to the spinal cord. There they synapse either with interneurons or with ventral horn motor neurons. Stimulations of the ventral horn neurons activates the skeletal muscles with which they are associated, The direct pathway primarily regulates fast and fine movements such as writing.
2.) The Indirect (Extrapyramidal) System: The indirect system includes brain stem motor nuclei and all motor pathways except the pyramidal pathways. These tracts were formerly lumped together as the extrapyramidal system because their nuclei of origin were presumed to be independent of the pyramidal tracts. This term is still widely used clinically. However, pyramidal tract neurons are now known to project to and influence the activity of most "extrapyramidal" nuclei, so modern anatomists prefer to use the term indirect, or multineuronal pathways.
These motor pathways are complex and multisynaptic. They are most involved in regulating the axial muscles that maintain balance and posture, the muscles controlling coarse limb movements, and head, neck, and eye movements that follow objects in the visual field. Many of the activities controlled by subcortical motor nuclei depend heavily on reflex activity.
Overall, the reticulospinal and vestibulospinal tracts maintain balance by varying tone of postural muscles. The rubrospinal tracts control flexor muscles, whereas the tectospinal tracts and the superior colliculi mediate head movements in response to visual stimuli.